
9.P/Q型カルシウムチャネルとシナプス刈込み
P/Q型カルシウムチャネル*1に着眼したキッカケは、7回目のコラムで紹介したGluD2の研究に端を発する。GluD2欠損マウスでは、平行線維が支配すべき遠位樹状突起にシナプス形成障害が起きて、そこに生じたフリースパインを標的として登上線維支配に遠位拡大が起こる(Kurihara et al., 1997; Ichikawa et al., 2001)。この現象から、平行線維と登上線維の2種類の興奮性入力によるシナプス形成は、互いに競合していることに気がついた。つまり、平行線維支配の強化分子であるGluD2がなくなると登上線維の支配が強まるならば、登上線維側にもその支配を強化する分子機構Xがあり、平行線維支配の過度な近位拡大を抑止しているはずである(図1)。運動会での綱引き競争に例えると、両チームの牽引力が釣り合うと綱の結節点は動かないが、一方の牽引力が弱くなれば結節点は牽引力の強い方へとズルズルと移動する。平行線維支配が遠位で登上線維支配が近位に分離しているという回路形態は、単に自然とそうなっているのではなく、競合という動的平衡の上に立って結果的にそうなっている。そう考えて、すぐに思い当たった分子機構Xの最有力候補が、P/Q型カルシウムチャネルである。
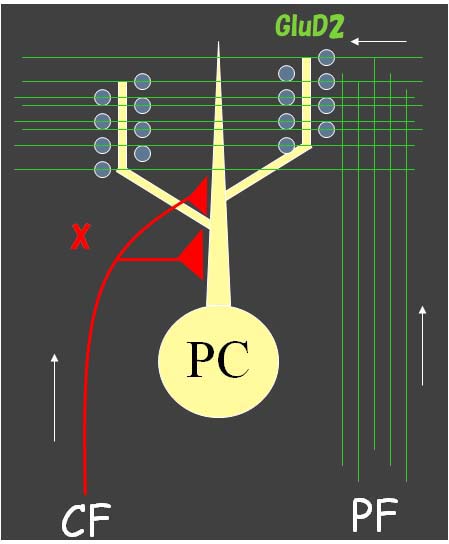
- 図1 登上線維支配の強化機構X
- 平行線維支配の強化機構であるGluD2が欠損すると、平行線維支配が弱まり、登上線維支配が遠位に向かって拡大した。同様に、登上線維支配の強化機構Xが欠失すれば、登上線維支配が弱まり、平行線維支配が近位に向かって拡大するであろうと、予測できる。(筆者作成図)
P/Q型カルシウムチャネルに着眼した理由
この分子に着目したのは、以下の3つの特性による。
第1に、このカルシウムチャネルはプルキンエ細胞に非常に豊富で、このニューロンのカルシウム電流の90%以上を占めている。そのため、P/Q型カルシウムチャネルの欠損は、この細胞のCa2+動態とそれに基づくニューロン機能を大きく変化しうる。
第2に、通常カルシウムチャネルは軸索の終末部に分布して、プレシナプス側からの神経伝達物質の放出に関わっている。ところが、P/Q型カルシウムチャネルはプルキンエ細胞の樹状突起にも豊富に分布して、細胞外からポストシナプス側へのCa2+流入も媒介する。一次視覚野や一次体性感覚野などの感覚性大脳皮質のシナプス回路発達において、視床皮質投射線維間の活動依存的なシナプス刈込みを駆動しているのが、NMDA型グルタミン酸受容体*2を介するCa2+流入である(Li et al., 1993; Kutsuwada et al., 1996)。一方、プルキンエ細胞は、発達初期をすぎるとNMDA受容体を持たなくなる例外的なニューロンである(Watanabe et al., 1993)。これらの状況証拠を考慮すると、P/Q型カルシウムチャネルがプルキンエ細胞のシナプス刈込みにおいてその役割を担っているのではないかと考えた。
第3に、このカルシウムチャネルは高閾値型*3に分類され、その活性化には強い脱分極性が必要である。1本の登上線維は、プルキンエ細胞の近位樹状突起に巻き付きながら数百もの興奮性シナプスを形成することから、単発の登上線維の活動により数百個もの登上線維シナプスからグルタミン酸が同時放出され、複合スパイク*4と呼ばれる活動電位をプルキンエ細胞に発生させる(Kano et al., 1992; Konnerth et al., 1992)。この強い脱分極作用が、平行線維に対抗する登上線維側の強化分子機構としてふさわしい、と考えた訳である。
そこで、P/Q型カルシウムチャネルのチャネル本体をコードするα1Aサブユニットのノックアウトマウスを韓国のShin教授から供与を受け、狩野方伸教授らと共同してP/Q型カルシウムチャネル欠損マウスの解析を開始した(図2)。この研究を担当したのが当時博士課程大学院生の宮崎太輔君(現、北海道大学保健学研究院准教授)であり、学位論文提出締切りの前日となる2003年12月に論文受理の連絡が届き、無事医学博士の学位も取得にもつながった(Miyazaki et al., 2004)。

- 図2 2004年4月に開催された米国コールド・スプリング・ハーバー研究所*5でのスナップショット
- 左から、宮崎太輔先生、Hee-Sup Shin教授(Korea Institute of Science and Technology)、筆者。この時、P/Q型カルシウムチャネルの論文報告を行い、感謝の気持ちをShin教授に直接伝えることができた。
登上線維支配の形態学的解析法の開発
P/Q型カルシウムチャネル欠損による表現型を紹介する前に、この研究のために宮崎先生が開発した登上線維支配様式可視化のための3重標識法を紹介したい(Miyazaki & Watanabe, 2011)。
20世紀における登上線維解析には生理学と解剖学の研究法があり、登上線維を選択的に可視化する解剖学研究法としては順行性神経トレーサー*6しかなかった。この方法では、まず登上線維の起始核である延髄下オリーブ核にトレーサーを注入し、数日後トレーサー標識された登上線維を小脳で観察する。先行して行っていたGluD2欠損マウスの登上線維支配の解析では、トレーサー標識された登上線維を千数百枚の連続電子顕微鏡観察法によち追跡することで、登上線維の多重支配を世界で初めて電顕レベルで示すことができた(Ichikawa et al., 2001)。
2000年に入り、グルタミン酸をシナプス小胞に充填する小胞膜グルタミン酸輸送体*7が3種類同定され(Ni et al., 1994; Aihara et al., 2000; Gras et al., 2002; Takamori et al., 2000)、このうちVGluT2が登上線維終末にVGluT1が平行線維終末に発現することから、免疫組織化学的に両者を識別することが可能になった(Miyazaki et al., 2003)。そこで、トレーサー標識登上線維の連続電顕に代わるより簡便な解析法として、VGluT2の蛍光標識/登上線維のトレーサー蛍光標識/プルキンエ細胞樹状突起の蛍光標識にトライした。この方法により、登上線維の多重支配と単一支配の判別と、多重支配における余剰な登上線維シナプスの同定が容易に行えるようになり(図3)、登上線維支配の新たな解剖学的解析法が確立した。
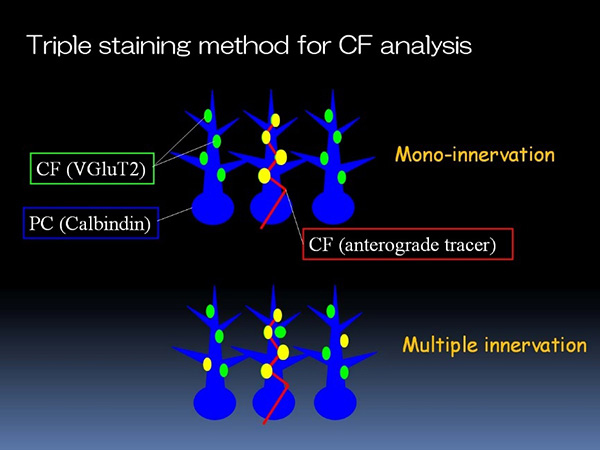
- 図3 VGluT2蛍光標識/トレーサー蛍光標識/プルキンエ細胞蛍光標識による新たな登上線維解析法
-
蛍光標識デキストランビオチン(赤)を下オリーブ核に投与し、一部の登上線維とその終末を赤色に蛍光標識する。数日後、小脳の固定切片を作成し、すべての登上線維終末をVGluT2抗体で緑色に、プルキンエ細胞の細胞体と樹状突起をカルビンジン*8抗体により青色に標識する。
単一支配であれば、トレーサー標識された登上線維により支配されるプルキンエ細胞上の登上線維終末は赤と緑の融合色である黄色に、トレーサー未標識の登上線維により支配されるプルキンエ細胞上の登上線維終末は緑色となって、細胞ごとに分離する(上段)。一方、多重支配であれば、黄色と緑色の登上線維終末が1個の細胞に混在し、その位置から余剰な登上線維支配の位置も特定できる(下段)。(筆者作成図)
P/Q型カルシウムチャネルとシナプス刈込み
P/Q型カルシウムチャネル欠損マウスでは、GluD2欠損マウスとは正反対の表現型、すなわち登上線維支配の近位退縮と平行線維支配の近位拡大を観察でき、予想は見事に的中した(Miyazaki et al., 2004)。
つまり、P/Q型カルシウムチャネル欠損マウスでは、登上線維による近位樹状突起の支配が弱まり、登上線維の分布範囲も分子層の半分程度にまで減少した。一方、近位樹状突起や細胞体から異所性のスパインが多数派出し、その多くをVGluT1陽性の平行線維終末が支配していた。前回のコラムにおいて、正常発生過程において平行線維シナプスはまず樹状突起の全域に形成され、生後15-20日の間に近位樹状突起から平行線維シナプスの刈込みが生じることを解説した。要するに、P/Q型カルシウムチャネル欠損マウスでは、mGluR1欠損マウスと同様にこの刈込みが障害される結果として、平行線維支配の近位拡大が生じたものと解釈することができる。
P/Q型カルシウムチャネル欠損マウスの小脳失調はきわめて重篤で、登上線維による多重支配の程度も電気生理学的解析では80%以上、解剖学的解析では全てのプルキンエ細胞において生じていた。3重染色による解析により、P/Q型カルシウムチャネル欠損マウスでは、プルキンエ細胞の近位樹状突起と細胞体において、トレーサー陽性/VGluT2陽性の登上線維とトレーサー陰性/VGluT2陽性の登上線維が混在して多重支配する様子を観察した(図4)。
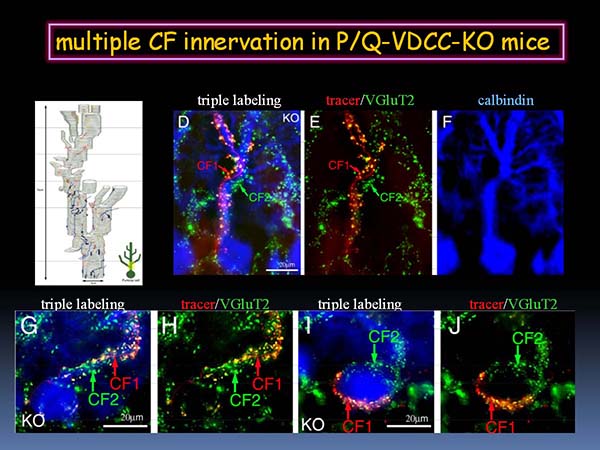
- 図4 P/Q型カルシウムチャネル欠損マウスにおける近位型の登上線維多重支配
- 黄色のトレーサー陽性/VGluT2陽性登上線維と、緑色のトレーサー陰性/VGluT2陽性の登上線維による多重支配が、近位樹状突起や細胞体で生じている。(筆者作成図)
これらの所見は、蛍光顕微鏡の画像のデコンボリューション処理による低解像度の画像で、解析したマウスも受精卵のときから遺伝子を欠損している完全欠失型遺伝子欠損マウスであるため、この表現型の原因の詳細は不明な点も多く残った。その後、三品教授や崎村教授らがプルキンエ細胞選択的な遺伝子欠損マウスを開発し、プルキンエ細胞選択的なP/Q型カルシウムチャネル欠損マウスにおいて、完全欠失型でみられたほとんどの表現型が再現され、プルキンエ細胞のポストシナプス側へのCa2+流入が活動依存的なシナプス刈込みを制御していることが明らかになった(Hashimoto et al., 2011; Miyazaki et al., 2012)。この間に顕微鏡も進歩していて、3波長レーザーによる共焦点レーザー顕微鏡による高精細な解析により証明することができた。現在、薬剤誘導によるプルキンエ細胞選択的な成体期にP/Q型カルシウムチャネル欠損マウスの解析も進めており、論文がまとまった段階で追加記載を行いたい。
前回のコラムと合わせ、mGluR1の活性化による細胞内ストアからのCa2+放出とP/Q型カルシウムチャネルの活性化による細胞外からのCa2+流入が、プルキンエ細胞における活動依存的なシナプス刈込みの駆動原理となっていることを解明できた。
文献
- Kurihara H, Hashimoto K, Kano M, Takayama C, Sakimura K, Mishina M, Inoue Y, Watanabe M. Impaired parallel fiber-Purkinje cell synapse stabilization during cerebellar development of mutant mice lacking the glutamate receptor δ2 subunit (GluRδ2). J Neurosci 17:9613-9623, 1997.
- Ichikawa R, Miyazaki T, Kano M, Hashikawa T, Tatsumi H, Sakimura K, Mishina M, Inoue Y, Watanabe M. Distal extension of climbing fiber territory and multiple innervation caused by aberrant wiring to adjacent spiny branchlets in cerebellar Purkinje cells lacking glutamate receptor d2. J Neurosci 22:8487-8503, 2002.
- Li Y, Erzurumlu RS, Chen C, Jhaveri S, Tonegawa S. Whisker-related neuronal patterns fail to develop in the trigeminal brainstem nuclei of NMDAR1 knockout mice. Cell 76:427-437, 1994.
- Kutsuwada T, Sakimura K, Manabe T, Takayama C, Katakura N, Kushiya E, Natsume R, Watanabe M, Inoue Y, Yagi T, Aizawa S, Arakawa M, Takahashi T, Nakamura Y, Mori H, Mishina M. Impairment of suckling response, trigeminal neuronal pattern formation, and hippocampal LTD in NMDA receptor e2 subunit mutant mice. Neuron 16:333-344, 1996.
- Watanabe M, Mishina M, Inoue Y. Distinct spatiotemporal expressions of five NMDA receptor channel subunit mRNAs in the cerebellum. J Comp Neurol 343:513-519, 1994.
- Kano M, Rexhausen U, Dreessen J, Konnerth A. Synaptic excitation produces a long-lasting rebound potentiation of inhibitory synaptic signals in cerebellar Purkinje cells. Nature 356:601-604, 1992.
- Konnerth A, Dreessen J, Augustine GJ. Brief dendritic calcium signals initiate long-lasting synaptic depression in cerebellar Purkinje cells. Proc Natl Acad Sci USA 89:7051-7055, 1992.
- Miyazaki T, Hashimoto K, Shin H-S, Kano M, Watanabe M: P/Q-type Ca2+ channel a1A regulates synaptic competition on developing cerebellar Purkinje cells. J Neurosci 24:1734-1743, 2004.
- Miyazaki T, Watanabe M. Development of an anatomical technique for visualizing the mode of climbing fiber innervation in Purkinje cells and its application to mutant mice lacking GluRδ2 and Ca(v)2.1. Anat Sci Int 86:10-18, 2011.
- Ni B, Rosteck PR Jr, Nadi NS, Paul SM. Cloning and expression of a cDNA encoding a brain-specific Na(+)-dependent inorganic phosphate cotransporter. Proc Natl Acad Sci USA 91:5607-5611, 1994.
- Aihara Y, Mashima H, Onda H, Hisano S, Kasuya H, Hori T, Yamada S, Tomura H, Yamada Y, Inoue I, Kojima I, Takeda J. Molecular cloning of a novel brain-type Na(+)-dependent inorganic phosphate cotransporter. J Neurochem 74:2622-2625, 2000.
- Gras C, Herzog E, Bellenchi GC, Bernard V, Ravassard P, Pohl M, Gasnier B, Giros B, El Mestikawy S. A third vesicular glutamate transporter expressed by cholinergic and serotoninergic neurons. J Neurosci 22:5442-5451, 2002.
- Takamori S, Rhee JS, Rosenmund C, Jahn R. Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neurons. Nature 407:189-194, 2000.
- Miyazaki T, Fukaya M, Shimizu H, Watanabe M. Subtype switching of vesicular glutamate transporters at parallel fibre-Purkinje cell synapses in developing mouse cerebellum. Eur J Neurosci 17:2563-2572, 2003.
- Hashimoto K, Tsujita M, Miyazaki T, Kitamura K, Yamazaki M, Shin HS, Watanabe M, Sakimura K, Kano M. Postsynaptic P/Q-type Ca2+ channel in Purkinje cell mediates synaptic competition and elimination in developing cerebellum. Proc Natl Acad Sci USA 108:9987-9992, 2011.
- Miyazaki T, Yamasaki M, Hashimoto K, Yamazaki M, Abe M, Usui H, Kano M, Sakimura K, Watanabe M: Cav2.1 in cerebellar Purkinje cells regulates competitive excitatory synaptic wiring, cell survival, and biochemical cerebellar compartment. J Neurosci 32:1311-1328,2012.
用語解説
- *1 P/Q型カルシウムチャネル P/Q-type calcium channel
- 膜電位の脱分極によって開口する電位依存性カルシウムチャネル(Voltage-dependent calcium channel, VDCC)の分子ファミリーは、チャネル本体を構成するα1サブユニットの種類により、L型、N型、P/Q型、R型、T型に分類される。このうち、Cav2.1遺伝子にコードされるαサブユニットがP/Q型カルシウムチャネルを構成する。P/Q型のPはプルキンエ細胞に由来する。
- *2 NMDA型グルタミン酸受容体 NMDA-type glutamate receptor
- イオンチャネル型グルタミン酸受容体のサブタイプの1つで、GluN1, GluN2A-D, GluN3A-Bサブユニットにより構成される。高いCa2+透過性と電位依存性Mg2+阻害として知られる特異なチャネル特性を有し、活動依存的の回路発達やシナプス可塑性の誘発に中心的役割を果たしている。
- *3 高閾値型カルシウムチャネル high-threshold VDCC
- カルシウムチャネルは、高い膜電位(~−20mV)で活性化するL型、N型、P/Q型、R型と低い膜電位(~−60mV)で活性するT型に大別される。
- *4 複合スパイク complex spike
- 登上線維からのシナプス入力は、樹状突起でのP/Q型カルシウムチャネルを介したCa2+流入を引き起こし、複数のピークを持つ特徴的な複雑スパイクをプルキンエ細胞に誘発する。これに対し、平行線維からのシナプス入力は、単一の波形となる単純スパイクsimple spikeを発生する。
- *5 コールド・スプリング・ハーバー研究所 Cold Spring Harbor Laboratory
- アメリカ合衆国ニューヨーク州ロングアイランドにある民間非営利財団による研究所で、生物学・医学の研究および教育を目的とする。学術誌や学術書の出版や、会議や講習会を開催している。
- *6 順行性神経トレーサー anterograde neuronal tracer
- 順行性神経トレーサーを投与すると、細胞体から取り込まれ、順行性軸索輸送により神経線維の末端部まで運ばれる。逆行性神経トレーサーは、神経線維の末端部で取り込まれ細胞体へと逆行輸送される。
- *7 小胞膜グルタミン酸輸送体 vesicular glutamate transporter
- 液胞型プロトンATPaseによって形成されるプロトン電気化学勾配を駆動力として、アミノ酸の1種であるグルタミン酸を細胞質からシナプス小胞内部に輸送する。当初無機リン酸を輸送する輸送体として同定されていたものが(Ni et a., 1994; Aihara et al., 2000)、2000年に小胞膜グルタミン酸輸送体であることが判明した(Takamori et al., 2000)。従来からのグルタミン酸作動性ニューロンはVGluT1とVGluT2を発現してグルタミン酸の小胞充填に関わるが、その発現部位や発現細胞は異なっていることが多い。一方、VGluT3は、従来から非グルタミン酸作動性と分類されてきたGABAニューロンやモノアミンニューロンに発現し、グルタミン酸と他の伝達物質の共放出に関わる。
- *8 カルビンジン calbindin
- 細胞質に分布するカルシウム結合タンパク質の1種で、特定のニューロンや細胞のマーカーとして広く用いられている。小脳では、プルキンエ細胞の選択的に発現する。